Частная генетика собакиГенетика окраскиИсследованием окраски собак занимались многие ученые. Сведения по генетике этого признака были опубликованы в монографиях Ильина (1932), Доусона (1937), Уитни (1947), Бернса и Фрезер (1966) и других. Многие авторы детально исследовали генетику окрасов собак одной породы.
В нашей стране экспериментальным изучением занимался в 30-е годы Н.А. Ильин в кинологической лаборатории и в Московском зоопарке.
Наиболее емким вкладом в проблему являются исследования К. Литтла, в течение многих лет изучавшего окрасы собак в Джексоновской лаборатории в США. Его работы шли в трех направлениях: экспериментальные скрещивания в лаборатории, анализ данных, полученных от заводчиков и владельцев питомников, племенная работа в своем питомнике. В течение 25 лет им было исследовано много тысяч пометов собак. Результатом явилась публикация монографии «The Inheritance of Coat Color in Dogs.» (1957).
Подробную сводку по окрасам собак приводит Р. Робинсон в монографии «Genetics for Dog Breaders» (1982).
Детальный анализ наследования окрасов собак дан в книгах М. Сотской «Пудель» (1998, 2004), Н. Московкиной и М. Сотской «Генетика и наследственные болезни собак и кошек» (2000).
Нужно отметить, что в работах разных авторов встречается несколько различающаяся символика в обозначении отдельных аллелей, а так же по-разному трактуются типы формирования того или иного окраса или места расположения отдельных аллелей в множественных сериях. Однако, в целом, разница во взглядах отдельных исследователей не носит принципиального характера.
Окрас шерстного покрова зависит от типа пигмента, формы пигментных гранул и распределения их по волосу. Все многообразие окрасок млекопитающих обусловлено наличием или отсутствием пигментов —
[b]меланинов[/b] .
Согласно современным представлениям меланин представлен двумя формами:
эумеланином и
феомеланином, которые существуют в виде пигментных зерен разнообразной формы. Восприятие цвета зависит от преломления света, проходящего через них или отражающегося от них. Эумеланин имеет две модификации:
черный и
коричневый. Феомеланин представлен
желтым и
красным вариантами.
Вложение:
 i_031.png [ 32.49 КБ | Просмотров: 1885 ]
i_031.png [ 32.49 КБ | Просмотров: 1885 ]
Вложение:
 i_032.png [ 40.01 КБ | Просмотров: 1885 ]
i_032.png [ 40.01 КБ | Просмотров: 1885 ]
Вложение:
 i_033.png [ 30.88 КБ | Просмотров: 1885 ]
i_033.png [ 30.88 КБ | Просмотров: 1885 ]
Вложение:
 i_034.png [ 8.63 КБ | Просмотров: 1885 ]
i_034.png [ 8.63 КБ | Просмотров: 1885 ]
Окраска собак Другие особенности экстерьера
Особенности экстерьера в основном являются полигенными и наследуются по типу промежуточного наследования, или как количественные признаки. При этом может проявляться как общая рецессивность, так и неполное доминирование. Многие из этих признаков проявляют неполную пенетрантность и экспрессивность. Некоторые сцеплены друг с другом. Поэтому менделевский анализ наследования морфологических особенностей в большинстве случаев оказывается затрудненным.
Строение тела собакиВажные породообразующие признаки, связанные с формой головы, теле и длиной конечностей, обусловлены особенностями синтеза в организме собаки гормона роста — соматотропина (СТГ) и чувствительности к нему тканей организма. Среди множества пород собак можно выделить три группы:
акромегалики , акромикрики и хондриодистрофики . Каждая имеет определенные фенотипические проявления динамического действия
СТГ.Акромегалики характеризуются вытянутыми в длину костями черепа и, частично, конечностей. Представителями этого типа являются различные борзые, колли и доберманы. Наследуется акромегалия по доминантному типу.
Акромикрики, наоборот, имеют укороченные кости лицевого черепа, так называемую мопсообразность, и, иногда, укороченные конечности. Эта группа представлена разными бульдогами, боксерами, пекинесами, мопсами, гриффонами и другими мопсообразными собаками. Выяснено, что акромикрия наследуется моногенно по рецессивному типу. Поэтому межпородные вязки приводят к потере типичного для этих пород облика.
При хондриодистрофии или иначе ахондроплазии задерживается рост трубчатых костей конечностей и развивается коротконогость, например, у таксы и вельшкорги. Подобные коротконогие собаки существовали еще в Древнем Египте и их изображения встречаются на древнеегипетских фресках. Хондриодистрофия также как акромикрия наследуется моногенно по рецессивному типу.
Коротконогость, возникающая за счет укорочения предплечий, по-видимому, имеет полигенную основу и может наследоваться по типу неполного доминирования. Аналогичным образом наследуется и укороченные бедро и голень, нарушающие правильный постав задних конечностей. Можно предполагать, что эти признаки имеют неполную пенетрантность и экспрессивность.
Исследования наиболее характерных для собак недостатков экстерьера показали, что многие признаки имеют отчетливо выраженный наследственный характер, однако могут усугубляться плохим выращиванием.
По-видимому, по доминантному типу наследуются такие признаки, как
укороченная спина и связанная с ней куполообразная поясница, укороченный и скошенный круп (короче говоря, нарушение соотношения 2:1:1). Показано, например,
что узкая грудная клетка доминирует над нормальной,
коротконогость, возможно не полностью, над
длинноногостью.
Отчетливо наследственный характер имеют
углы сочленений.
Рост и вес тела собак представляют типичный пример количественных признаков. Как количественный признак наследуется и размер
черепа.
Относительно же формы головы, как показывает практика,
длинномордость доминирует над
короткомордостью,
широкий череп и
нижняя челюсть над
узкими,
короткая теменная часть — над длинной. Метисы собак акромегалических пород (колли, борзые) имеют столь же длинную лицевую часть черепа, однако более широкую мозговую часть. Данный эффект сохраняется и в последующих поколениях.
Доминантным признаком с неполной пенетрантностью и экспрессивностью являются часто встречающиеся у собак
прибылые пальцы на задних ногах.
Врожденное укорочение и дефекты хвоста могут быть обусловлены как доминантными, так и рецессивными генами, что подтверждается многочисленными наблюдениями. Гибриды бульдогов с собаками, имеющими хвосты нормальной формы, обычно обладают хвостами нормальной длины и формы.
Некоторые авторы указывают, что хвост саблевидной формы у собак доминирует над крючкообразным, кольцевидным и хвостом-поленом.
Часто, при скрещиваниях собак разных пород наблюдается появление признаков характерных для дикого типа, так называемый, уход в
дикий тип. Это может быть объяснено неаллельностью генов, определяющих фенотип данных животных, а также наличием специфических регуляторных элементов генома. Частичные возвраты в дикий тип возможны и при скрещивании животных одной породы, происходящих из географически разобщенных популяций.
Зубная системаСтроение зубной системы является исключительно важным элементом экстерьера собак. Значительные отклонения в форме, количестве зубов и прикусе может свидетельствовать о нарушениях генетической конституции не только отдельного животного, но и всей линии или семейства, к которому оно относится. Аномалии зубной системы наряду с другими аномалиями, например крипторхизмом, могут рассматриваться как маркеры гибридного дисгенеза.
История собаководства и многочисленные исследования селекционеров показывают, что в значительной мере наследственно обусловлена
форма прикуса . Наследственно обусловлены врожденное укорочение нижней челюсти или
прогнатия и укорочение верхней челюсти —
прогения, также приводящие к образованию неправильного прикуса.
Большое внимание селекционеров привлекает также проблема полнозубости. Для некоторых пород наличие полного комплекта зубов является абсолютным. Для других — требования несколько смягчены. Врожденные аномалии количества зубов такие как
олигодонтия и
полиодонтия могут быть диагностированы как в момент актирования помета, так и после смены зубов. Генетическая обусловленность этих аномалий не вызывает сомнений.
Врожденные отклонения от нормального числа премоляров, их размеров и формы обнаруживаются у всех представителей отряда хищных. Можно выделить следующие механизмы их возникновения: 1) расщепление зубного зачатка; 2) слияние зубных зачатков; 3) развитие дополнительного зубного зачатка в зубной пластинке; 4) отсутствие одного или нескольких зачатков в зубной пластинке.
Часть недостатков зубной системы оказываются связанными с излишним удлиннением лицевой части черепа, как, например, у доберманов. Этот процесс часто идет преимущественно за счет верхней челюсти и сопровождается формированием укороченной или слишком мелкой нижней челюсти. Возрастание длины челюстей приводит к появлению лишних премоляров и моляров. При слишком узкой морде, резцовый ряд нижней челюсти чрезмерно изгибается вперед. Это приводит к тому, что при недостатке места закладываются только пять или даже четыре резца. Подобное явление отмечается и у собак мелких пород, имеющих относительно крупные зубы. Замечено, что у пород, имеющих голову классического строения, например у немецких овчарок, обнаруживается меньшее количество дефектов резцов.
Резцовая олигодонтия может проявляться по разному:• зубы нормального размера, но количество их меньше — четыре или пять вместо положенных шести;
• два зуба срастаются вместе от корней до верха коронок образуя один более крупный зуб нормальной формы. Если это происходит с одной стороны, то вырастает пять зубов, а если с двух — то четыре;
• два крайних зуба срастаются своими корнями, коронки зубов оказываются разделенными своими верхушками. Образуется, так называемый сдвоенный зуб.
Вложение:
 i_035.png [ 7.67 КБ | Просмотров: 1885 ]
i_035.png [ 7.67 КБ | Просмотров: 1885 ]
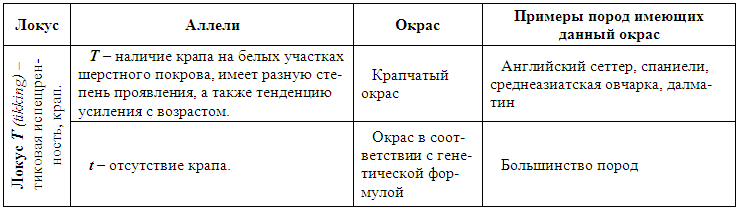
Рис. 21. Олигодонтия по резцам и премолярам: 1 — челюсти с нормальным количеством резцов; 2 — пять резцов в верхней челюсти, второй зуб слева сдвоенный, образован путем срастания коронок; 3 — челюсти с уменьшенным количеством резцов: пять резцов в верхней челюсти и четыре в нижней; 4 — пять резцов в нижней челюсти. Крайний правый зуб образован в результате срастания корней двух соседних; 5 — нормальный резец; 6 — сдвоенный резец, образованный срастанием коронок и частично корней; 7 — сдвоенный резец, образованный срастанием корней двух соседних зубов; 8 — олигодонтия по премолярам нижней челюсти. Отсутствует Р1; 9 — олигодонтия по премолярам верхней челюсти. Отсутствует Р2Многозубость или иначе
[b]полиодонтия[/b] встречается в форме типичной, или атавистической, для которой характерно наличие избыточных зубов внутри зубного ряда и которая бывает почти физиологичной и атипичной, когда сверхкомплектные зубы вырастают вне зубных лунок, иногда даже вне ротовой полости.
Псевдополиодонтия чаще всего связана с сохранением молочных зубов. Сверхкомплектные зубы могут быть представлены в виде сдвоенного зуба.
Вложение:
 i_036.png [ 5.79 КБ | Просмотров: 1885 ]
i_036.png [ 5.79 КБ | Просмотров: 1885 ]
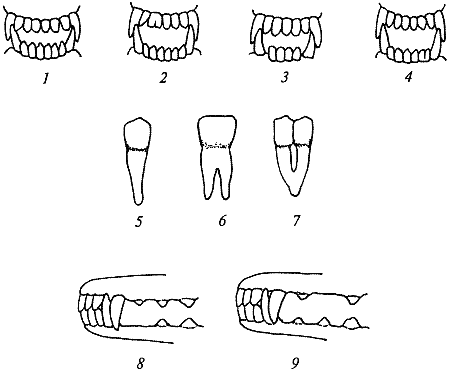
Рис. 22. Полиодонтия: 1 — по резцам; 2 — по клыкам, образованная невыпавшим молочным клыком
Полиодонтия часто являются следствием недостаточности щитовидной железы. Некоторые породы имеют повышенную частоту полиодонтии по резцам.
Иногда у собак карликовых и той-пород зубы бывают слишком крупными по отношению к челюсти и не могут расположиться в ровную линию, что приводит к возникновению, так называемой,
нерядности. Нерядность может быть связана, также, и с недоразвитием нижней челюсти. Как показывает опыт, степень наследственной обусловленности нерядности достаточно высока.
В некоторых случаях встречается, так называемый,
перекос, проявляющийся в наклоне зубного ряда в определенную сторону. Большинство заводчиков и владельцев собак склонны объяснять это травмой или склонностью собаки грызть кость или игрушку одной стороной челюсти, а также регулярным потягиванием поводка или палки в игре. Однако, как показывают генетические исследования, данный дефект или предрасположенность к нему имеют определенную наследственную детерминированность.
У породных собак, особенно мелких пород, достаточно часто наблюдаются отклонения в прорезывании зубов. Например, постоянные зубы прорезываются при невыпавших молочных, и образуется
двойной комплект зубов, чаще всего клыков, или даже, полный
двойной ряд зубов .
Вложение:
 i_037.png [ 4.96 КБ | Просмотров: 1885 ]
i_037.png [ 4.96 КБ | Просмотров: 1885 ]
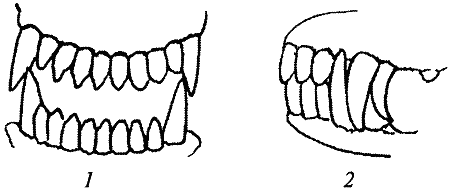
Рис. 23. Нерядность резцов и перекос челюстей: 1 — нерядность резцов нижней челюсти; 2, 3 — перекос челюстейПодобные аномалии характерны для нарушений деятельности системы гипофиз — щитовидная — паращитовидная железа, что также встречается при несбалансированности генома. Иногда при смене молочных зубов задерживается появление премоляров, и, реже, моляров и, несмотря, на героические усилия владельца щенка, зубы все же не появляются и рентгенограмма показывает, что не были заложены даже их зачатки. Нельзя говорить о моногенном или полигенном наследовании этих нарушений, скорее этот факт говорит о том, что зубная система в процессе формирования чрезвычайно чувствительна как к повреждающим моментам, так и к несогласованной деятельности различных частей генома. В подтверждение теории несбалансированности говорит тот факт, что часто нарушения зубной системы возникают при скрещиваниях собак определенных линий.
При детальном исследовании зубной системы иногда обнаруживается ненормальное сближение корней зубов —
конвергенция корней . При этой аномалии не происходит рассеяния давления производимого корнями зубов на ткани челюсти, а наоборот, возникает сильное локальное давление, со временем приводящее к трофическим нарушениям в деснах и окружающих тканях.
Дивергенция корней, или иначе расхождение коронок, возникает в период смены молочных зубов в результате задержки их выпадения и приводит к развитию редкозубости. Довольно часто встречается
ретенция зуба — при этом зуб находится не в зубном ряду, а в толще челюстной кости.
Таким образом, форма прикуса и всякие отклонения от полной зубной формулы в большинстве случаев определяются полигенной системой, что может быть рассмотрено как показатель неудачной комбинации блоков генов в генотипе, способной повлечь за собой множественные нежелательные последствия. Носителей этих аномалий желательно исключать из разведения.
Форма ушейДлительная селекционная работа, проводимая на собаках, привела к формированию множества форм ушей. Можно выделить следующие три группы ушей: стоячие, висячие и полустоячие.
Стоячие уши направлены концами вперед и вверх. Соразмерно голове собаки стоячие уши могут быть
большими или
малыми. Уши могут иметь более или менее заостренные концы, быть близкими по форме к равнобедренному треугольнику с основанием короче боковых сторон, или напоминать равносторонний треугольник. Концы стоячих ушей могут быть направлены в разные стороны, образуя
развешенные или
сближенные уши.
Полустоячие уши имеют крепкие хрящи только в нижней половине уха, вторая же половина, в силу мягкости хряща, опускается вниз или в сторону.
Висячие уши бывают двух видов:
висячие на хряще с крепким у основания хрящом, поддерживающем ухо на линии лба (ухо эрдель-терьеров) и
висячие — хрящи которых мягки и уши, в силу своей тяжести, свисают по обеим сторонам головы собаки (у различных пород гончих и легавых). Висячие уши также могут быть длинными и короткими, а также узкими и широкими с закругленными или острыми концами.
Независимо от постава, уши могут быть
высоко и
низко посаженными.
Велико разнообразие и в степени
оброслости ушных раковин шерстью. Стоячие уши обычно покрыты короткой прямой шерстью; висячие могут быть покрыты как короткой, так и длинной шерстью.
Наследование формы и размера ушной раковины полигенно, но можно выделить определенные локусы, оказывающие значительное влияние на отдельные детали ее строения.
Так показано, что при скрещивании собак с висячими ушами с собаками со стоячими ушами, наблюдается неполное доминирование со стороны висячего уха.
HH — висячее ухо;
hh — стоячее ухо;
Hh — полустоячее ухо.
В этом же локусе присутствует и аллель доминирующий полностью над
H и
h — Hа — полустоячее ухо типа «колли». При этом собаки с генотипами
HаHа, HаH и
Hаh фенотипически неразличимы. Таким образом, в популяциях колли, терьеров и других пород, могут присутствовать все три аллеля, что приводит к периодическому выщеплению собак со стоячими ушами.
Очевидно, такой тип наследования характерен для внутрипородных популяций. В случае, если ушные раковины скрещиваемых пород сильно различаются по размеру, эта закономерность может быть утеряна.
Также показано неполное доминирование и взаимозависимость размеров ушей. Если висячее ухо маленькое, то доминирует стоячее ухо, но получившееся стоячее ухо больше, чем у исходной формы. Если висячее ухо большое или длинное, то стоячее ухо доминирует не полностью. Но, тем не менее, гибриды F1, полученные от скрещивания пуделей с волками и шакалами («пуво» и «пуша»), имели стоячие уши. Гибриды F2имели уши разнообразной формы и размера.
Авторы наблюдали целый ряд межпородных гибридов и проследили наследование нескольких признаков. Гибриды F1пуделя и английского бульдога (с ушами розовидной формы) имели уши по форме и размеру похожие на пуделиные, однако более высоко посаженные. Гибрид F1пуделя и фокстерьера имел полустоячие уши в типе керриблютерьера. Гибриды F1карело-финской лайки и фокстерьера имели стоячие уши. Гибриды F1эрдельтерьера и ризеншнауцера имели уши по типу близкие к ушам ризеншнауцера.
Окраска глаз собакиОкраску глаз собак можно условно разделить на две категории: нормальную и аномальную.
Нормальная окраска глаз собаки зависит от локусов окраски шерсти
Е,В,D и может быть более или менее темной — темно-карей или черной, ореховой или карей и желтой. Эти окраски соответствуют трем аллелям
Jr; Jrm и iry, которые, по-видимому, не полностью доминируют одна над другой. У собак с ослабленной пигментацией, гомозиготных по рецессивному аллелю
dd — цвет глаз дымчатый, что мы иногда наблюдаем у серорожденных пуделей и догов. У рецессивных гомозигот
bb (коричневых) при генотипе
Jr — цвет глаз каштановый, при
irm — светло-ореховый и
iry — светло-желтый.
При
В — Jr — дает почти черный цвет глаз,
irm — темно карий,
iry — карий.
К
аномальным окраскам глаз можно отнести рубиновоглазие, гетерохромию радужной оболочки и альбиностические. Подобные окраски глаз не представляют собой патологии, и, чаще всего, не оказывают влияния на остроту зрения собак.
Рубиновоглазие, проявляется в том, что при освещении под определенным углом в глазах собаки появляется рубиново-красный отсвет. Этот феномен не зависит от цвета глаз. Исследования показали, что у таких собак наблюдается своеобразное расположение пигмента в радужной оболочке глаза, которое не влияет на зрение собаки и передается как рецессивный признак. Аллель определяющий его развитие обозначен как
р. Отмечены случаи сочетания рубиновоглазия со светло-серым окрасом шерсти у пекинесов.
У альбиносов полностью отсутствует пигмент в радужной оболочке глаза и, в связи с тем, что через ее толщину просвечивают сосуды она имеет красный цвет.
Альбинизм глаз различной степени отмечался среди фокстерьеров, мальтийских болонок, боксеров, догов и далматинов. Но в целом альбиносы среди собак весьма редки.
Гетерохромия радужной оболочки («беловзор», «сорочий глаз») связана с аномалией радужной оболочки, которая у животных состоит из трех слоев различной толщины: переднего и заднего и лежащей между ними стромы. В нормальном глазу строма более или менее пигментирована за счет насыщенности пигментными клетками, также за счет пигментного эпителия заднего слоя радужной оболочки, что и определяет естественный цвет глаз. Этот самый задний слой у «сорочьего глаза», в отличие от альбиностического нормально пигментирован. Лежащие впереди его пигментные клетки стромы имеют веретенообразную форму, они сильно разветвлены, заполнены пигментом и снаружи уплотняются, образуя густую сеть — пограничный слой. Лишь недостаток пигмента в строме радужной оболочки ведет к клиническому проявлению гетерохромии радужной оболочки, называемой «беловзор», или «сорочий глаз». Полное отсутствие пигментных клеток стромы обуславливает полную и частичную гетерохромию радужки. Из-за большой рыхлости соединительной ткани глаза радужная оболочка «сорочьего глаза» выглядит голубоватой, в связи с этим точнее его было бы назвать «галочьим». Как при полной, так и неполной гетерохромии радужки, участки, лишенные пигмента, обычно отличаются фарфоровой белизной с серо-голубой зоной зрачка и краем склеры; но и здесь при сужении зрачка серо-голубой оттенок усиливается.
В зависимости от локализации и формы различают:
1) одностороннюю частичную гетерохромию радужки (правостороннюю или левостороннюю);
2) одностороннюю полную гетерохромию радужки (правостороннюю или левостороннюю);
3) двустороннюю частичную гетерохромию радужки;
4) левостороннюю полную гетерохромию радужки с правосторонней частичной или наоборот;
5) двустороннюю полную гетерохромию радужки;
Почти половина всех случаев связана с редукцией пигмента в вентральной части глаза, примерно, 30%, — в медиальной, и лишь относительно редко встречаются участки дорсальной, латеральной или циркулярной локализации.
Как показало большинство исследований, представляется, что гетерохромия не оказывает никакого влияния на остроту зрения и светочувствительность. Такой глаз во всех случаях реагирует на свет непосредственно обычным резким сужением зрачка. В сумерках зрачок выглядит совершенно нормально. У собак описаны случаи, когда с гетерохромией радужки нередко сочетались другие тяжелые нарушения глаз, как микрофтальмия и колобома, а также глухота.
У различных пород собак гетерохромия радужки сочетается с пятнистым или мраморным окрасом. Однако, между этими признаками не существует абсолютной корреляции. Гетерохромия наследуется обычно как простой аутосомно-рецессивный признак, однако некоторые авторы описывают его как аутосомно-доминантный.
Шерстный покровВ процессе породообразования шерстный покров собак претерпел значительные изменения. Среди них можно встретить как шерсть близкую к «дикому» типу, разные варианты мягкой, жесткой и полужесткой, длинной и совсем короткой и, даже, практически голых.
Волосы собак могут иметь разнообразную форму.
Прямой волос имеет прямой, лишь с небольшим изгибом в месте расширения стержень; изогнутый — с постепенным изгибом в одну сторону: волос
с надломом — с резким переломом в одну сторону ;
волнистый — имеет стержень, отклоняющийся от прямой оси в обе стороны;
кольцеобразный или
спиральный — закручен в одну сторону и образует плотные кольца или спирали.
Часть пород сохраняет
«дикий» или скорее
«нормальный» тип шерсти: двухъярусный мех и неодинаковую длину и фактуру шерсти на разных частях тела.
У собак и диких собачьих можно выделить, по крайней мере, три разновидности шерсти дикого типа:
1) «Нормальный» — шерсть средней длины. Присутствует у большинства видов средней полосы — волков, шакалов, лесных и дальневосточных кошек и у многих пород естественного облика: лаек, овчарок, гончих средней полосы и т. д.
2) «Укороченная шерсть дикого типа» — у динго и новогвинейской собаки и оцелота среди диких форм. У собак укороченную шерсть дикого типа имеют мопсы, стаффордширские терьеры, бульдоги и т. д.
3) «Удлиненная шерсть дикого типа» — встречается в природе у лисиц, песцов, енотовидных собак, красных волков. Среди домашних собак у самоедов, вольфшпицев, кесхаундов, чау-чау, колли, кавказских овчарок и т. п.
Всю массу короткошерстных собак можно разделить на собак с укороченной шерстью дикого типа с выраженным подшерстком и волосами всех типов и гладкошерстных собак. Гладкошерстные собаки обычно не имеют подшерстка, при этом покровные волосы в ряде случаев имеют незначительные отличия на разных частях тела. У некоторых собак эти различия практически нивелируются.
Очевидно, гладкошерстность может быть определена иными аллелями, чем короткошерстность дикого типа. Короткошерстность дикого типа является доминантной по отношению к длинной шерсти, а гладкошерстность — рецессивной.
Исследователями описан ряд аллелей, определяющих некоторые свойства шерстного покрова собак. Однако необходимо отметить, что их обозначения и трактовка разными авторами не всегда совпадают.
Длина шерсти обозначается символом
L.L — шерсть «дикого» типа; l 1— длинная шелковистая шерсть; l2 — гладкая короткая шерсть. По-видимому, доминирование в этом локусе имеет следующий порядок: L› l 1 › l2.
Аллель l1 продлевает период роста волоса, что и способствует увеличению его длины. Возможно, что вариации длины шерсти и особенностей строения определяются рядом генов-модификаторов данного аллеля.
Длинношерстные породы могут сохранять выраженные отличия покровных волос и подшерстка (колли, ретриверы, ньюфаундленды и т. д.) или почти не иметь подшерстка как, например, сеттеры. У некоторых из них остевые волосы стали настолько тонкими, что практически не отличаются от подшерстка (йоркширские терьеры, афганы). У собак типа пуделей, командоров и пули остевые волосы и подшерсток имеют одинаковую длину и перевиваются друг с другом.
Распределение волос разной длины по корпусу у длинношерстных собак также имеет разный характер. Некоторые из них сохраняют дикий тип оброслости с длинной шерстью на теле, хвосте и на бедрах и укороченной на морде и ногах (колли, шпицы). Другие имеют
равномерную оброслость по корпусу, ногам и морде (комондоры, пудели, мягкошерстные терьеры, ши тцу), так называемую
брудастость. При скрещивании собак этих двух типов доминирующим оказывается второй тип.
Очевидно, можно предположить наличие особого локуса, ответственного за характер оброслости. Мы предлагаем обозначить данный локус как W (wild) и выделить, по меньшей мере, два аллеля W — равномерную оброслость по всему телу собаки и w — обусловливающий дикий тип оброслости.
Курчавая шерсть может быть также очень разной: волосы могут закручиваться в спираль, или кольца, быть похожими на металлическую стружку или равномерно волнистыми. Возможно, что курчавая шерсть ирландских водяных спаниелей и ретриверов, а также волнистая шерсть кокеров и других пород может определяться разными генами.
Скрещивание пуделей с кокерами и другими породами с подобной шерстью дает в первом поколении собак с шерстью типа кокера. Это показывает, что данные аллеи относятся в одному локусу и волнистая шерсть доминирует над скрученной.
Таким образом, можно унифицировать обозначения данных аллелей следующим образом: wa — неволнистая шерсть, wa1 — волнистая шерсть , wa2 — скрученная (спиральная) шерсть. Доминирование аллелей в этом локусе очевидно носит следующий характер: wa›wa1› wa2.
Универсальным для всех жесткошерстных пород собак является ген жесткошерстности Wh. Рецессивный аллель этого локуса обусловливает развитие шерсти дикого типа. При генотипе llWh — волосы длиннее, чем при LLWh-, но короче, чем при llwhwh. Порядок доминирования аллелей в этом локусе: Wh›wh.
Шерсть такого типа встречается у собак с жесткой прямой шерстью — шнауцеров, брюссельского гриффона, некоторых терьеров, жесткошерстной таксы, дратхаара. Метисы собак этих пород большей частью также имеют шерсть данного типа. Многие курчавошерстные терьеры при скрещивании с прямошерстными породами полностью утрачивают завиток и их метисы выглядят так же, как и метисы собак с жесткой прямой шерстью.
Жесткие волосы часто бывают извитыми или курчавыми, поэтому иногда бывает трудно провести четкую грань между жесткошерстными, полужесткошерстными и курчавыми собаками.
Курчавая, скрученная и волнистая шерсть в сочетании с аллелями Wh и wh и L и l, определяют шерстный покров целого ряда пород. Пудели, очевидно, имеют генотип Wh-llkk или WhWhllkk. При скрещивании их с волками или собаками с диким типом шерсти получаются собаки с довольно длинной прямой и достаточно жесткой шерстью, скрученность, курчавость или волнистость шерсти полностью исчезает. Метисы различных спаниелей, полученные при скрещивании с длинношерстными собаками, имеют весьма характерный фенотип и шерсть близкую по типу к спаниелям. Метисы спаниелей и короткошерстных собак имеют обычно шерсть дикого типа.
Мы предлагаем следующую систему формирования шерстного покрова собак (табл. 10).
Кроме вышеописанных аллелей, обусловливающих развитие тех или иных характеристик шерстного покрова существует масса факторов, как наследственного, так и ненаследственного характера, также оказывающих влияние на формирование типа шерсти у животного.
К наследственным можно отнести множество генов-модификаторов, влияющих на развитие и соотношение волос разного типа (подшерстка и остевых волос), а также возможную неполную пенетрантность и экспрессивность описанных аллелей, вкупе обеспечивающих большую широту нормы реакции, что, в свою очередь, способствует многообразию проявлений модификационной изменчивости шерстного покрова.
К ненаследственным можно отнести влияние климатических условий, тип кормления, наличие или отсутствие стрессорных факторов и т. д. Даже внутри одной породы могут встречаться собаки с разной шерстью. Многое зависит и от способов ухода за ней. Так, например, у одного и того же эрдельтерьера после стрижки или щипки сформируется совершенно разный волосяной покров, отличающийся и по структуре и, даже, по интенсивности окраски.
Вложение:
 i_038.png [ 17.74 КБ | Просмотров: 1885 ]
i_038.png [ 17.74 КБ | Просмотров: 1885 ]
Таблица 10. Система формирования шерстного покрова собак
У собак существует целый ряд мутаций, приводящих к развитию
безволосости и отмеченных в разных точках земного шара. Встречаются разные обозначения безволосости собак. Так Н.А. Ильин (1932) обозначает эти аллели следующим образом: N — безволосость; n — нормальное состояние шерстного покрова. Робинсон обозначает ген безволосости как Hr , и hr как нормальное состояние шерстного покрова. Аллели безволосости в гомозиготном состоянии приводит к внутриутробной гибели его носителей. Все живущие голые собаки являются гетерозиготными и при скрещивании между собой дают в каждом помете гомозиготных особей, имеющих нормальный шерстный покров, так называемых «пуховок».
У мексиканских и китайских голых собак редкий шерстный покров есть только на голове и ногах, на теле присутствуют лишь отдельные волосы, возможно типа вибриссов. Кожа голых собак испещрена рисунком мягких складок, увеличивающих истинную поверхность кожи и выполняющих множественные защитные функции.
Сама по себе их кожа достаточно тонкая, слой подкожной жировой клетчатки также невелик. Отсутствие волос и сравнительно тонкая кожа, очевидно не могут обеспечить достаточно хорошую термоизоляцию, поэтому голые собаки легко мерзнут и нуждаются в предохранении от охлаждения.
Безусловно аллели безволосости, как и аллели дикого типа, обладают
плейотропным действием. Сам факт внутриутробной гибели щенков, гомозиготных по гену безволосости, говорит о глубоком нарушении органогенеза. Плейотропное действие гена безволосости проявляется и в формировании определенного породного фенотипа. В частности зубы у голых собак по своему строению во многом отличаются от зубов собак других пород.
Очевидно, бесшерстность китайских и мексиканских голых собак обусловлена разными аллелями. Экспериментальные вязки мексиканских голых собак с беспородными собаками, подходящего размера и фенотипа (за исключением бесшерстности) приводят к рождению в пометах голых щенков по фенотипу не отличимых от породных, в то же время, среди пуховок наблюдается огромный разброс по фенотипу. Пуховки мексиканских голых собак полученные в породных пометах также весьма разнообразны и не допускаются к выставкам и племенному разведению.
Китайские голые собаки генетически более стабильны и их пуховки имеют определенный фенотип, что позволяет им участвовать в выставках и племенной деятельности.
Кроме доминантного гена безволосости с плейотропным действием у собак встречается и рецессивный ген безволосости, обусловливающий развитие шерстного покрова у американских голых терьеров. Щенки американского голого терьера рождаются опушенными и теряют шерсть к 6–8 неделям жизни. Интересно отметить, что собаки этой породы не имеют аномалий зубной системы, типичных для других голых собак.
Очень интересная особенность шерстного покрова встречается
у риджбеков. У собак этой породы на спине имеется гребень, образованный волосами, растущими в направлении, противоположном росту остальных волос, так называемый «ридж». Иногда гребней бывает два. Стандарт породы требует, чтобы гребни были четко обозначены, симметричны и сужались к хвосту. Этот признак наследуется рецессивно и обозначается символом ds.
Как показывают исследования, проведенные на разных видах животных, оказывается довольно эффективной массовая селекция по длине или густоте шерсти. Однако между ними существует отрицательная корреляция, так как высокая частота расположения волосяных фолликулов уменьшает площадь их питания, что, и регламентирует рост шерсти в длину. Подобную корреляцию можно отметить и у собак — длинная шерсть йоркширских терьеров, ши тцу, два типа шерстного покрова встречается и у пуделей: густой с хорошо выраженным подшерстком и медленно растущий, не достигающий значительной длины и более редкий, но отличающийся большей длиной.
Рост шерсти также зависит от физиологического состояния организма, уровня кормления и значительно замедляется во время беременности и лактации.



